
Fucoidan, a complex sugar molecule from brown seaweed rich in fucose and sulfates, has demonstrated diverse biological effects. Its anti-cancer properties have been noted in numerous investigations, shown by its power to cause apoptosis and stop cell division in laboratory and in vivo models.
This blog post will introduce a study titled “Pathway Analysis of Fucoidan Activity Using a Yeast Gene Deletion Library Screen,” authored by Monika Corban et al. The objective of this research was to pinpoint cellular pathways influenced by fucoidan through a non-biased genetic methodology, to uncover novel understandings regarding fucoidan’s anti-cancer properties.
The study aimed for an unbiased perspective on gene-fucoidan interactions by assessing the growth of a comprehensive collection of 4,733 S. cerevisiae haploid gene deletion strains, with and without the addition of 500 μg/mL wakame fucoidan (UPF). The drug-gene interactions of wakame fucoidan were assessed by systematic screening of the entire library of 4,733 haploid S. cerevisiae gene deletion strains. The findings were partially corroborated through cell cycle analysis and DNA damage detection conducted on non-immortalized human skin fibroblasts and colon cancer cells.
The research selected a single, thoroughly studied cell line to test the application of yeast gene pathways in mammalian systems. The general toxicity of UPF was assessed in HCT-116 colorectal cancer cells to confirm UPF-induced cell cycle regulation and DNA damage in mammalian cells.
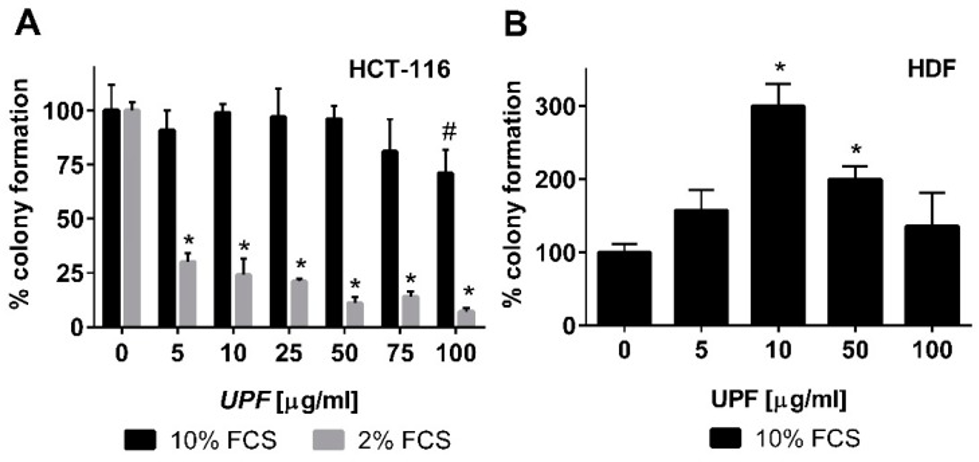
To assess the immediate impact of fucoidan therapy, HCT-116 colon cancer cells in their logarithmic growth phase were subjected to UPF for more than 24 hours. Subsequently, WST-1 conversion was quantified. Regardless of the serum concentration in the cell culture medium, UPF concentrations up to 100 µg/mL showed no signs of toxicity to colon cancer cells in the WST-1 assay. In fact, UPF concentrations as low as 2.5 µM appeared to significantly increase WST-1 dye conversion. This effect probably stems from the redox-active properties of the UPF formulation, so we will examine cell viability in its presence through other tests. Therefore, in contrast to the WST-1 assay, which only detects acute toxicity of compounds based on metabolic activity, the colony formation assay was employed to also assess the long-term effects of UPF treatment. Consistent with the WST-1 data, UPF had little effect on colony numbers after two weeks of continuous treatment. Under the most favorable cell culture conditions, as depicted in Figure 1A, it was observed that only the most concentrated UPF solution, specifically at 100 µM, resulted in a statistically significant reduction of roughly 25% in the number of colonies, with a p-value less than 0.05.
With the serum concentration in the cell culture medium decreased to 2%, the experiment was repeated for a more detailed evaluation of this effect. Growth factor depletion is a commonly used approach to increase the stringency of this assay and detect mild, non-lethal toxicity. This reduction in FCS content did not alter overall cell cloning efficiency in the absence of UPF (colony numbers in untreated cultures at 2% and 10% FCS were similar), but it had a dramatic effect on UPF activity. A notable decrease in colony count, around 70%, was observed even at the lowest UPF concentration, with statistical significance (p < 0.001), as shown in Figure 1A. As depicted in Figure 1A, this toxicity escalated in a dose-dependent fashion, leaving only 7% of colonies viable at the maximum UPF concentration.
Next, to determine if this toxicity is unique to immortalized cells or affects other cell types, the researchers investigated the impact of UPF on the colony-forming ability of non-immortalized primary human dermal fibroblasts (HDFs). These cells are remarkably sensitive to culture conditions, forming colonies only in at least 10% fetal calf serum (FCS). Therefore, lower FCS concentrations could not be tested. In contrast to HCT-116 cells, UPF treatment of HDF cells significantly increased the number of colonies between 10 and 50 µM, demonstrating a bell-shaped concentration curve. As shown in Figure 1B, HDFs exhibited colony formation at least equal to untreated controls when exposed to the highest UPF concentration of 100 µM.
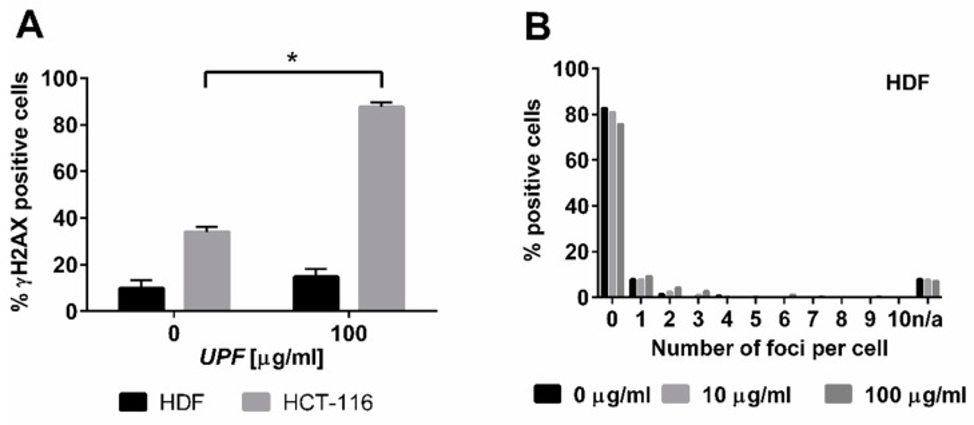
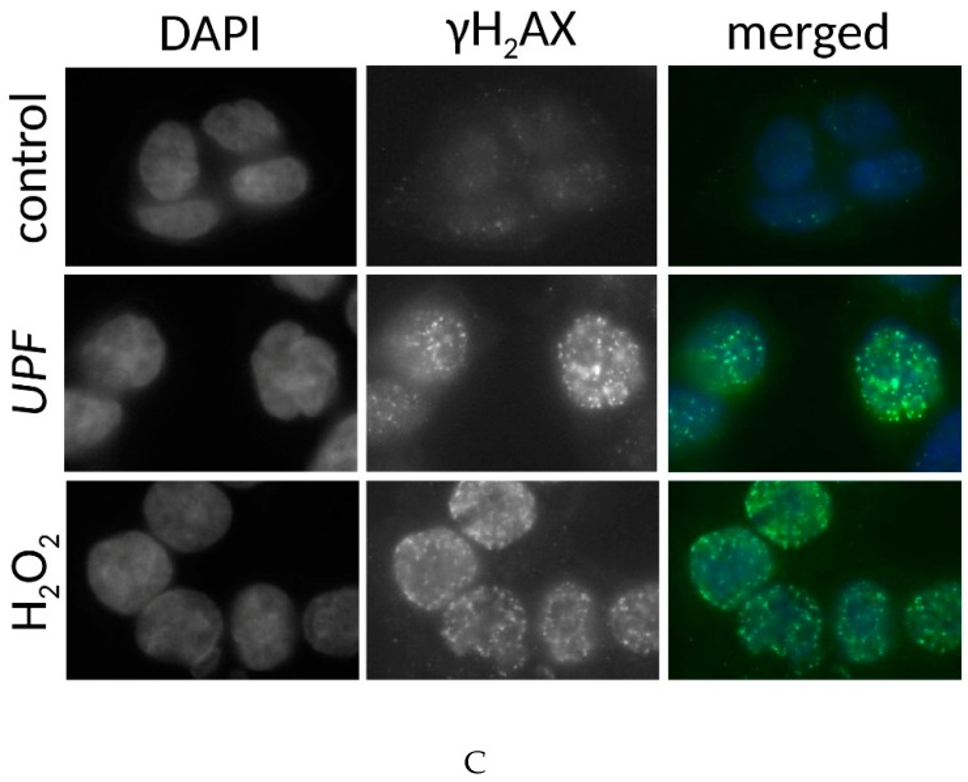
Data from yeast experiments show a similar toxicity in colon cancer cell lines, implying that UPF interacts with DNA repair mechanisms. The rationale is that DNA damage induction is the common modus operandi for many anticancer drugs. Nuclear γH2AX staining is a marker of DNA damage-induced cellular repair activity. The investigation into UPF’s potential to induce DNA damage involved exposing non-immortalized human dermal fibroblasts (HDFs) and colon cancer cells (HCT-116) to 100 µg/mL UPF for more than 24 hours. In untreated HDFs, only approximately 10% of cells displayed γH2AX foci, and these cells typically displayed fewer than four foci.
Following UPF exposure, there was no observable rise in the quantity of positive cells, nor did these positive cells display a greater number of foci, as depicted in Figures 2A and 2B. In HDFs displaying γH2AX foci, the count of these foci was at its lowest point, as illustrated in Figure 2B. In contrast to HDFs, approximately 40% of HCT-116 cells harbored γH2AX foci in the absence of UPF, indicating genetic instability characteristic of many tumor cell lines. Moreover, HCT-116 cells showed a highly significant (p < 0.01) UPF-induced effect, with approximately 90% of them containing γH2AX foci after 24 hours, as shown in Figure 2A. In contrast to HDFs, HCT-116 cells also exhibited significantly higher numbers of γH2AX foci per cell after UPF exposure as shown in Figure 2C.
Cell cycle analysis via flow cytometry was performed to clarify the chronic toxicity of UPF on HCT-116 cells as observed in the colony formation assay. UPF induced a significant (p < 0.01) G1 arrest after 72 hours, but no significant difference was observed at earlier points. The reduction in S-phase cells approached significance (p = 0.052), but the number of G2-phase cells was unaffected.
To summarize, 292 genes potentially involved in the cellular response to wakame fucoidan extract (UPF) were identified by exposing a deletion library of S. cerevisiae to it. The pathway analysis categorized these genes into various pathways and cellular functions, such as ribosome function and biogenesis, cell cycle signaling and DNA repair, nucleotide and amino acid biosynthesis, peroxisome biogenesis, mitochondrial function and energy metabolism, RNA synthesis, protein synthesis, and transport. Two typical pathways were identified in the human colon cancer cell line HCT-116. In this cell line, UPF suppressed colony formation, slowly induced G1 arrest, and caused significant DNA damage. This was not observed in non-immortalized primary human skin fibroblasts. The presented results showcase fucoidan’s multifaceted effects on distinct cellular processes within eukaryotic cells and contribute to a more profound understanding of how wakame fucoidan impedes the proliferation of human cancer cells.
Source: Mar Drugs. 2019 Jan 14;17(1):54. doi: 10.3390/md17010054