
A significant health problem globally is the overconsumption of salt, which is prevalent in nearly all countries. A high-salt diet (HSD) is strongly correlated with many diseases, including hypertension, chronic kidney disease, and autoimmune diseases. One of the primary ways in which a diet high in salt exacerbates the initiation and advancement of HSD-related illnesses is through its pro-inflammatory actions. Despite this, there is an incomplete understanding of how salt affects inflammatory responses in the body.
This blog post will discuss the study “L-fucose and fucoidan alleviate high-salt diet-promoted acute inflammation” by Wenhua Li et al. The research demonstrated that consuming a high-salt diet (HSD) lowers L-fucose, a gut bacteria metabolite, and intensifies inflammatory reactions in mice.
A standard acute peritonitis model using zymosan A was established to investigate the pro-inflammatory actions of a high-sugar diet (HSD). Mice were fed either a control diet (CD) or a high-sugar diet (HSD) for eight weeks. Following this period, zymosan A was administered intraperitoneally (i.p.), and flow cytometry was performed to measure the effects of the HSD.
The findings indicate that HSD plays a dual role in peritonitis, not only attracting inflammatory cells but also boosting the release of inflammatory cytokines. TNF-α and IL-6 are commonly used markers to assess inflammation, and MCP-1 is a chemokine that recruits circulating leukocytes to the site of inflammation. The combined results strongly imply that HSD aggravates the inflammatory process in mice suffering from zymosan A-induced peritonitis.
Following the confirmation of HSD’s pro-inflammatory action in zymosan A-induced peritonitis, the study then delved into the mechanisms responsible for this effect. Measurement of Na+ concentration in peritoneal fluid supernatant at different time points revealed no significant difference in Na+ concentration between the HSD and control groups in biochemical analysis. This suggests that the inflammation caused by HSD does not involve an increase in Na+ in the peritoneal cavity. Additionally, to examine if HSD contributes to inflammation indirectly by influencing the gut microbiome, mice in both the HSD and control groups were treated with broad-spectrum antibiotics for four weeks to eliminate gut bacteria.
The researchers examined how antibiotics affected the peritonitis response. They observed that after administering antibiotics, inflammatory cytokine levels, such as TNF-α, were elevated in both HSD and control groups relative to those not treated with antibiotics. However, there were no significant differences between the HSD and control groups in total cell count, neutrophil and monocyte/macrophage infiltration, or the expression of major inflammatory cytokines such as TNF-α, MCP-1, and IL-6 at 6 and 12 hours after zymosan A injection. These results strongly suggest that HSD-induced inflammation is likely mediated through abnormalities in the gut microbiota.
The study aimed to investigate the influence of a high-fat diet on the gut microbiota by analyzing the diversity and composition of mouse fecal microbiomes with 16S rRNA gene sequencing. In fecal samples collected from both groups, we found 8 to 12 phyla, 34 to 48 families, and 40 to 54 genera. After 8 weeks of HSD feeding, a decrease in the abundance and diversity of fecal bacteria was observed, although this decrease was not statistically significant based on α-diversity. The Jaccard index analysis further highlighted a marked distinction between the groups, underscoring a significant transformation in the gut microbiota’s composition after the consumption of HSD.
Taxonomic composition analysis also revealed significant changes in the composition of the gut bacteria. The most prevalent phyla at this taxonomic level were Bacteroidetes and Firmicutes. Moreover, Firmicutes were found to be abundant in the HSD group, while Bacteroidetes were abundant in the control diet (CD) group. There was a statistically significant shift, with a decrease in Bacteroidetes and an increase in Firmicutes, after the consumption of HSD. To identify representative bacteria associated with HSD, LEfSe analysis (LDA score (log10)>2) was performed, identifying 47 types of bacteria from phylum to genus level. In the CD group, researchers observed abundant Bacteroidetes, while the HSD group showed an abundance of Firmicutes. The cumulative findings indicate that an eight-week period of high-sucrose diet intake results in a reduction in both the quantity and variety of gut microorganisms, along with a substantial change in the microbial community structure.
Next, they aimed to identify gut metabolites that might contribute to the increased inflammation promoted by HSD. After excluding unidentified metabolites, HSD was found to downregulate eight specific gut bacterial metabolites: 1-methylhistamine, betaine, D-mannitol, deoxycytidine, methylmalonic acid, prephenic acid, L-fucose, and taurolithocholic acid. Of these, only L-fucose and betaine have been reported to have anti-inflammatory effects. However, dietary betaine also serves as a substrate that is metabolized by bacteria into trimethylamine and possibly trimethylamine N-oxide, which are associated with an increased risk of stroke, heart attack, and death. This led the researchers to undertake a further investigation into the mechanisms by which L-fucose promotes HSD-induced inflammation.
L-fucose is typically found in glycans and is released from these molecules by gut bacteria via an enzyme called fucosidase. The released L-fucose is then further metabolized and utilized in the body. First, they examined the effect of HSD on the abundance of fucosidase genes in gut bacteria by comparing the two groups using phylogenetic investigation of communities by reconstruction of unobserved states (PICRUSt2) and found that the abundance of fucosidase genes was significantly reduced in the HSD group.
Furthermore, they analyzed the relationship between genus-level bacterial communities identified by LEfSe and metabolites identified in negative ion mode using Spearman’s rank correlation coefficient. The outcome indicated a significant link between L-fucose levels and a greater presence of bacterial communities, specifically exhibiting a negative correlation with dominant bacterial communities in the HSD group and a positive one with the dominant bacterial communities in the CD group. This correlation suggests that these microorganisms are involved in the metabolism of L-fucose. They hypothesized that an insufficient amount of fucosidase in the gut’s microorganisms results in lower L-fucose production among the HSD individuals.
An in vivo study was performed to examine the anti-inflammatory properties of L-fucose. Peritonitis was induced in mice by administering zymosan A. The findings, illustrated in Figure 5A, revealed a substantial decrease in the total cell count, neutrophils, and monocytes/macrophages within the peritoneal cavity of mice treated with both HSD and L-fucose. Furthermore, the expression levels of inflammatory markers such as TNF-α, MCP-1, and IL-6 in the peritoneal fluid were also decreased. Next, the effects of L-fucose on RAW264.7 cells and peritoneal macrophages (PM) were evaluated. The study’s quantitative PCR findings demonstrated a concentration-dependent reduction in Mcp1 and Il6 gene expression in RAW264.7 cells upon treatment with L-fucose. Similarly, the relative expression levels of Mcp1 and Il6 were also reduced in PM. These results indicate that L-fucose can alleviate inflammation both in vivo and in vitro.
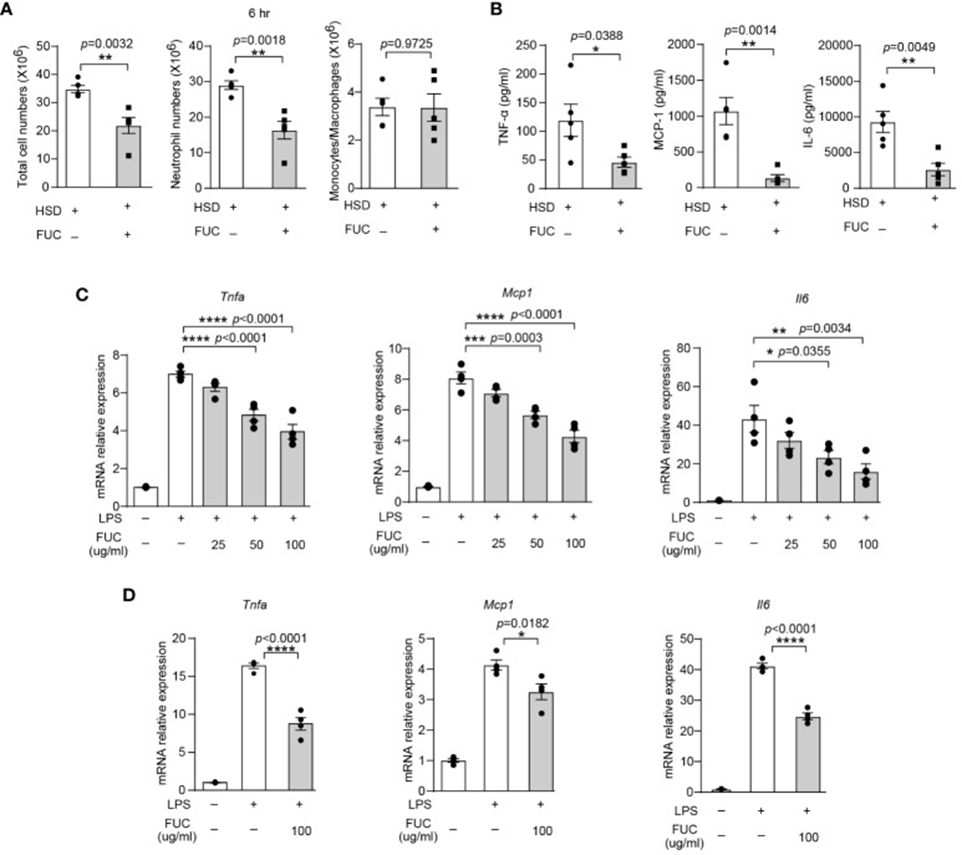
The data above revealed that L-fucose can alleviate inflammation induced by a high-fat diet (HFD). This discovery prompted speculation about whether fucoidan could exhibit anti-inflammatory properties comparable to L-fucose. To investigate whether fucoidan could ameliorate the exacerbation of inflammation caused by HFD, cells and cytokines in the peritoneal fluid of both the fucoidan-treated group and the HFD group six hours after zymosan A injection were analyzed by flow cytometry. As depicted in Figure 1A, the fucoidan-treated group exhibited lower quantities of both total cells and neutrophils in their peritoneal fluid compared to the HFD group. Furthermore, the levels of inflammatory cytokines TNF-α, MCP-1, and IL-6 in the supernatant of the FUC group were decreased compared to the HFD group, as per Figure 1B.
Additionally, in vitro experiments involving RAW264.7 cells and peritoneal macrophages (PM) were performed, and these experiments’ outcomes provided additional support for the findings. The relative mRNA expression levels of Tnfa (RAW264.7 cells; PM) and Mcp1 were decreased by fucoidan treatment. Figure 1C and D illustrate a decrease in IL-6 expression. These findings indicate that fucoidan, much like its primary constituent L-fucose, exhibits anti-inflammatory properties and can mitigate inflammation that is worsened by a high-fat diet.
This research indicated that eating a diet high in sugar (HSD) leads to a decrease in the variety and quantity of gut bacteria. It also changes the makeup and byproducts of these bacteria, which are vital for the progression of peritonitis. Furthermore, the findings indicated that lower L-fucose concentrations in the gut lead to more severe acute inflammation in mice when subjected to HSD. Based on the data obtained, it is suggested that L-fucose and fucoidan could play a role in suppressing the amplification of inflammatory processes initiated by HSD.
Source: Front Immunol. 2024 Mar 26;15:1333848. doi: 10.3389/fimmu.2024.1333848