
Malignant glioma (MG) is a type of brain tumor that has a bleak outlook, as it is prone to recurrence even after comprehensive treatment, resulting in a 5-year survival rate of only around 20%. The search for safer and more effective treatments is urgently needed. In multiple studies, it has been proven that Fucoidan can effectively induce the differentiation of osteoblasts in both human osteoblasts and adipose-derived stem cells. Additionally, in humans, Fucoidan has been found to stimulate osteoblasts through the activation of c-Jun N-terminal kinase (JNK) and extracellular signal-related kinase (ERK)-dependent bone morphogenetic protein 2 (BMP2)-Smad 1/5/8 signaling pathways. Furthermore, there have been reports of it stimulating the process of differentiation.
In this blog, I would like to share the study, “Epigenetic Modification and Differentiation Induction of Malignant Glioma Cells by Oligo-Fucoidan” by Chien-Huang Liao et al. In their study, the researchers focused on investigating the impact of wakame-derived Oligo-Fucoidan (OF) on the concept being studied in MG cells, specifically grade III U87MG cells (glioma cell) and grade IV glioblastoma multiforme (GBM) 8401 cells. They also examined the effect of wakame-derived Oligo-Fucoidan (OF) on immortalized astrocytes. The cells were compared with SVGp12 cells. The Oligo-Fucoidan (OF) used in this study is a low molecular weight (<667 Da) Fucoidan and is derived from the glycolytic degradation products of the original Fucoidan from the brown alga Laminaria japonica.
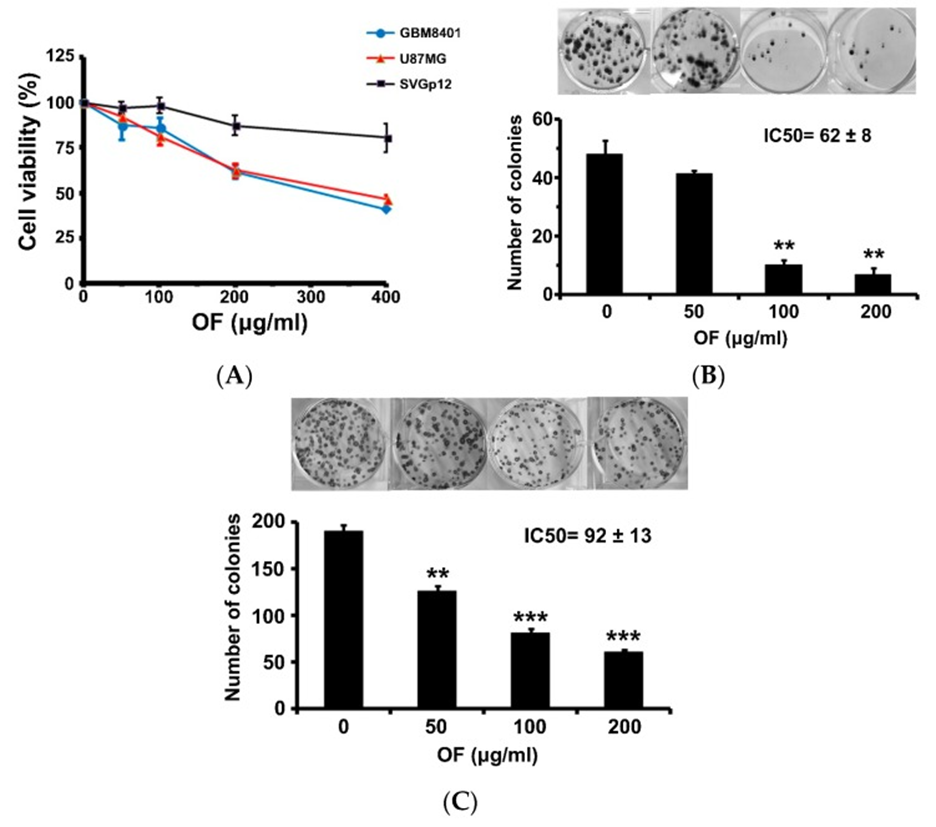
The proliferation of human MG cells (GBM8401 and U87MG) was measured using the sulforhodamine (SRB) assay after 72 hours of exposure to OF. The results showed a significant growth inhibition. In Figure 1A, it can be observed that cell proliferation of GBM8401 and U87MG cells was significantly reduced to 40% and 46% of the control at a concentration of 400 μg/mL. However, OF only had a minimal impact on the growth of immortalized astrocyte SVGp12 cells at the same concentration, indicating that OF primarily targets cancer cells. When the concentration of OF was at 200 μg/mL, it had a significant impact on colony formation of GBM8401 and U87MG cells, reducing it to 14% and 32% respectively as shown in Figures 1B and C. The 50% inhibitory concentration (IC50) of OF on clonogenicity of GBM8401 and U87MG cells upon 12 days of treatment was 62 ± 8 and 92 ± 13 μg/mL, respectively as shown in Figure. 1B and C. Higher-grade MG cells appear to be more sensitive to OF.
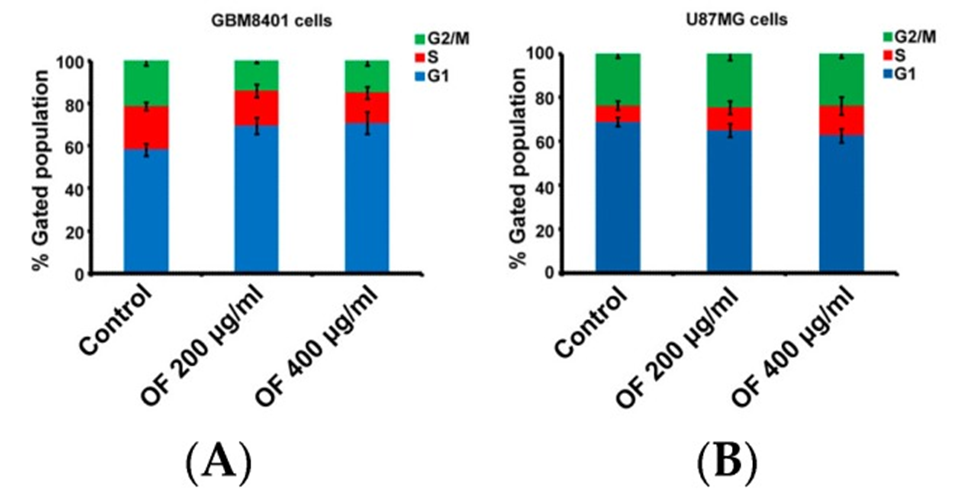
The cell cycle of GBM8401 cells was effectively halted by OF, as evidenced by the increase in the proportion of the G1 phase from 58% (control) to 69% and 71%, respectively (as depicted in Figure. 2A). In U87MG cells, the S phase increased from 7% (control) to 10% and 14% depending on OF concentration. (See Figure 2B) The result obtained from this study indicates that OF has the ability to inhibit cell proliferation in various types of MG cells by causing cell cycle arrest in either the G1 or S phase.
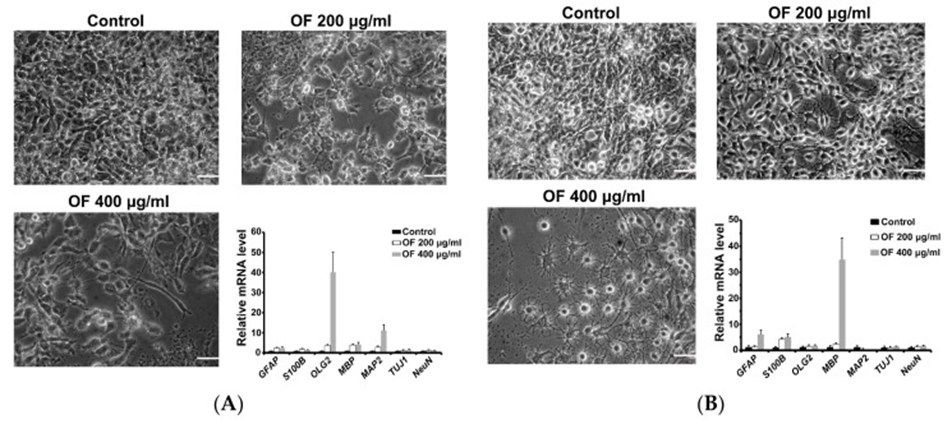
Next, early (astrocytes (GFAP), Oligodendrocytes (Olig2) and neurons (MAP2 and Tuj1)) and terminal (astrocytes (S-100β), Oligodendrocytes (myelin basic protein, MBP) and neurons (NeuN )) and measured differentiation markers in OF-treated MG cells by quantitative PCR assay. As shown in Figure 3A, neuronal and Oligodendrocyte-like cell shapes were observed in OF-treated GBM8401 cells. These GBM8401 cells showed a significant increase in early Oligodendrocyte (Olig2) and neuronal (MAP2) differentiation markers, as shown in Figure 3A, which supports the argument. In OF (400 μg/mL)-treated U87MG cells, a cell shape with many Oligodendrocyte-like cells and fewer glial cells was observed. (See Figure 3B) As a result, it was observed that the U87MG cells exhibited a substantial increase in the expression of the terminal Oligodendrocyte differentiation marker MBP, along with a significant upregulation of astrocyte markers (GFAP and S100B), as seen in Figure 3B. The joint action of OF can lead to the redifferentiation of MG cells, which in turn triggers malignant transformation through the dedifferentiation phenomenon.
They investigated the molecular mechanisms underlying the differentiation of OF-treated MG cells. the previous study found that OF can induce miR-29b and suppress DNMTs (DNMT1, 3A, and 3B) in cancer cells, and they hypothesized that DNMTs in MG cells could induce differentiation by OF. Consistent with expectations, OF decreased the protein levels of DNMTs in both GBM8401 and U87MG cells. The involvement of epigenetic mechanisms, specifically DNA demethylation, in the induction of differentiation by OF suggests that they may have a significant role to play.
The substantial decrease in DNMT3B protein levels caused by OF in U87MG cells may lead to a reduction in the methylation of the p21 gene and a potential restoration of its expression. In U87MG cells, the simultaneous use of the clinical DNMT inhibitor decitabine and OF can result in a synergistic effect, leading to the inhibition of proliferation and induction of MBP. Following the confirmation of its effectiveness in living organisms, it is necessary to conduct appropriate clinical trials to assess the potential of this complementary approach to treating MG.
Source: Mar Drugs. 2019 Sep; 17(9): 525. doi: 10.3390/md17090525